Estructura de los virus
Los virus presentan una amplia diversidad de morfologías, es decir, de formas y tamaños. Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1400 nm, sin embargo, solo miden unos 80 nm de diámetro. La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas. Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como el wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.
|
Retroviridae. |
Myoviridae. |
Cystoviridae. |
|
Virgaviridae. |
Lipothrixviridae. |
Inoviridae. |
|
Ampullaviridae. |
Fuselloviridae. |
Spiraviridae. |
|
Nimaviridae. |
Barnaviridae. |
Pithoviridae. |
|
Rhabdoviridae. |
Ascoviridae. |
Clavaviridae. |
|
Reoviridae. |
Alphasatellitidae. |
Adenoviridae. |
|
Baculoviridae. |
Poxviridae. |
Endornaviridae. |
|
Parvoviridae. |
Tombusviridae. |
Rudiviridae. |
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros. Algunos virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped (virus con envoltorio), mientras que otros carecen de ella (virus desnudos). La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica. Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside. En general, hay cuatro tipos principales de morfología vírica:
- Helicoidal
- Las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, que pueden ser cortos y muy rígidos o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.
- Icosaédrica
- La mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de estos también se componen de otros seis capsómeros y reciben el nombre de hexones.
- Con envoltura
- Algunas especies de virus tienen una envoltura, que es una forma modificada de una de las membranas celulares, ya sea la membrana externa que rodea una célula huésped infectada o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltura víricae. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, y la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de los virus que afectan a los animales poseen envoltorio y dependen de él para infectar.100
- Complejos
- Los virus tienen una cápside que no es ni puramente helicoidal, ni puramente icosaédrica, y que puede poseer estructuras adicionales como colas proteicas o una pared exterior compleja. Algunos bacteriófagos (como el Fago T4) tienen una estructura compleja que consiste en un cuerpo icosaédrico unido a una cola helicoidal (esta cola actúa como una jeringa molecular, atacando e inyectando el genoma del virus a la célula huésped), que puede tener una base hexagonal con fibras caudales proteicas que sobresalgan.
Los poxvirus son virus grandes y complejos con una morfología inusual. El genoma vírico está asociado con proteínas dentro de una estructura discal central conocida como nucleoide. El nucleoide está rodeado por una membrana y dos cuerpos laterales de función desconocida. El virus tiene una envoltura exterior con una espesa capa de proteína en la superficie. La partícula en general es ligeramente pleomorfa, con una forma que puede ir de la de un huevo a la de un ladrillo. El mimivirus es uno de los tipos de virus más grandes conocidos, con un diámetro en su cápside de 400 nm. De su superficie se proyectan filamentos proteicos de 100 nm. La cápside, vista al microscopio electrónico, tiene una forma hexagonal, de manera que probablemente es icosaédrica.
Algunos virus que infectan a las arqueas tienen estructuras inusuales, que no están relacionadas con ningún otro virus conocido. De igual manera, algunos bacteriófagos pueden tener diferentes estructuras en cuanto a su cola, con formas algo raras con respecto a otros virus.
Los virus con cápsides icosaedricas son estructuralmente similares a los microcompartimientos bacterianos y los nanocompartimientos de encapsulina, un conjunto de orgánulos proteicos encontrados en las bacterias y arqueas, que desempeñan funciones metabólicas y nutricionales.
Genoma
| Propiedad | Parámetros |
|---|---|
| Ácido nucleico |
|
| Forma |
|
| Cadenas |
|
| Sentido |
|
Se puede ver una enorme variedad de estructuras genómicas entre las especies de virus que, como grupo, contienen una diversidad genómica superior a la de los reinos de las plantas, los animales o las bacterias. Hay millones de diferentes tipos de virus; y únicamente alrededor de 5000 de ellos han sido descritos detalladamente. Los virus tienen un genoma compuesto de ADN o bien de ARN, y reciben respectivamente los nombres de «virus ADN» y «virus ARN». La gran mayoría de virus utilizan el ARN. Los virus de las plantas tienden a tener ARN monocatenario y los bacteriófagos tienden a tener ADN bicatenario.
Los genomas víricos pueden ser circulares, como los polyomaviridae o lineales, como los adenoviridae. El tipo de ácido nucleico es irrelevante para la forma del genoma. En los virus ARN, el genoma a menudo está dividido en partes separadas dentro del virión, y se le califica de «segmentado». Cada segmento suele codificar una proteína y los segmentos suelen estar reunidos en una cápside. No es necesario que cada segmento se encuentre en el mismo virión porque el virus en general es infeccioso, como lo demuestra el Brome mosaic virus.
Poco importa el tipo de ácido nucleico, un genoma vírico puede ser o bien monocatenario o bicatenario. Los genomas monocatenarios consisten en un ácido nucleico no emparejado, similar a una de las mitades de una escalera de mano cortada a lo largo por la mitad. Los virus bicatenarios consisten en dos ácidos nucleicos emparejados y complementarios, similares a una escalera de mano entera. Algunos virus, como los Hepadnaviridae, contienen un genoma que es parcialmente bicatenario y parcialmente monocatenario.

Bacteriófagos de la familia Cystoviridae infectando dos bacterias.
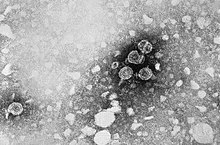
Micrografía mostrando viriones de hepatitis B.

Virófago infectando al Mamavirus.
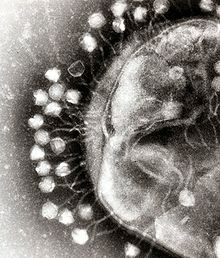
Bacteriófagos de los Caudovirales infectando una bacteria.

Bacteriófago perteneciente al orden de los Caudovirales.
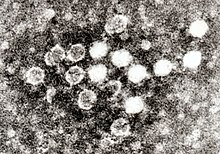
Parvovirus, cada virión mide 20-30 nm. Este virus es perteneciente al virus ADN monocatenario.

Micrografía electrónica de un rotavirus. La barra mide 100 nm.
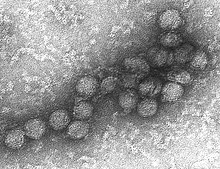
Virus del Nilo Occidental perteneciente al orden Flaviviridae.
Micrografía del virus de la inmunodeficiencia humana.

Micrografía hecha del virus de Ébola.
En los virus ARN o los virus ADN monocatenarios, las cadenas pueden ser o bien positivas (cadenas plus) o negativas (cadenas minus), dependiendo de si son complementarias en el ARN mensajero (ARNm) vírico. El ARN viral positivo es idéntico al ARNm viral y por tanto puede ser traducido inmediatamente por la célula huésped. El ARN viral negativo es complementario del ARNm y por tanto debe ser convertido en ARN positivo por una ARN polimerasa antes de ser traducido. La nomenclatura del ADN es similar a la del ARN, en cuanto a la «cadena codificadora» del ARNm vírico que le es complementaria (–), y la «cadena no codificadora» que es una copia (+).
El tamaño del genoma varía mucho entre especies. Los genomas víricos más pequeños solo codifican cuatro proteínas y pesan unos 106 daltons; los más grandes pesan unos 108 daltons y codifican más de un centenar de proteínas. Los virus ARN suelen tener genomas más pequeños que los virus ADN debido a una tasa de error más alta a la hora de replicarse, y tienen un límite superior de tamaño. Por encima de este límite, los errores en la replicación del genoma hacen que el virus sea inofensivo o incluso, incompetente. Para compensar esto, los virus ARN a menudo inician un proceso de segmentación en el que el genoma es separado en moléculas más pequeñas, reduciendo así las posibilidades de error. En cambio, los virus ADN tienen genomas mayores gracias a la elevada fidelidad de sus enzimas de replicación.
Los virus sufren cambios genéticos por diversos mecanismos. Entre estos se incluye un proceso llamado deriva genética en el que las bases individuales del ADN o el ARN mutan en otras bases. La mayoría de estas mutaciones puntuales son imperceptibles, pues la proteína que codifica el gen no cambia, pero aun así, pueden conferir ventajas evolutivas como resistencia a los medicamentos antivíricos. El cambio antigénico se produce cuando hay un cambio significativo en el genoma del virus. Esto ocurre como resultado de una recombinación genética. Cuando esto se produce en los virus de la gripe pueden ocurrir pandemias. Los virus ARN suelen existir como quasiespecies o en enjambres de virus de la misma especie, pero con secuencias de nucleósidos del genoma ligeramente diferentes. Estos grupos son un objetivo destacado por la selección natural.
Los genomas segmentados ofrecen ventajas evolutivas; diferentes cepas de un virus con el genoma segmentado pueden intercambiar y combinar genes, produciendo virus progenénicos (o descendientes) con características únicas. Esto recibe el nombre de «sexo vírico».
La recombinación genética es el proceso por el cual una cadena de ADN se rompe y luego se une al extremo de una molécula de ADN diferente. Esto se puede producir cuando diferentes virus infectan las mismas células al mismo tiempo, y estudios de la evolución de los virus han demostrado que la recombinación tiene un papel muy importante en las especies estudiadas.112 La recombinación es común en los virus ARN y ADN.
Ciclo replicativo de los virus

Ciclo replicativo de los virus.

Representación esquemática del proceso de replicación de un flevovirus.
- La adhesión o adsorción es una unión específica entre proteínas de la cápside vírica y receptores específicos de la superficie celular del huésped, pero algunos bacteriófagos también son capaces de adherirse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedante. Para que esto suceda la bacteria debe contener el factor sexual «F» o ciertas colicinas (factores de resistencia contra agentes antimicrobianos). Los bacteriófagos filamentosos con ADN de cadena sencilla se adhieren a las puntas de estos pili mientras que los bacteriófagos esféricos de ARN se adhieren a los costados de estos. La especificidad de unión —proteína y cápside— se determina por la variedad de huéspedes de los virus. Por ejemplo, el VIH solo infecta linfocitos T humanos, pues su proteína de superficie, gp120, puede interactuar con la CD4 y con receptores de la superficie del linfocito T. Este mecanismo ha evolucionado para favorecer los virus que solo pueden infectar células en que se pueden replicar. La adhesión al receptor que puede inducir cambios en la proteína de la envoltura viral que resultan en la fusión de las membranas viral y celular.
- La penetración sigue a la adhesión; los virus se introducen en la célula huésped mediante endocitosis mediada por receptores (también llamado viropexis) o por fusión de membrana. Esto recibe a menudo el nombre de penetración vírica. La infección de las células vegetales es diferente a la de las células animales. Las plantas tienen una pared celular rígida hecha de celulosa y los virus solo pueden entrar en las células cuando se produce un trauma en la pared celular. Los virus como el virus del mosaico del tabaco también pueden moverse directamente a las plantas, entre células, a través de poros llamados plasmodesmos. Las bacterias, como las de las plantas, tienen una fuerte pared celular que los virus tienen que romper para infectar la célula. Algunos virus han evolucionado mecanismos para inyectar su genoma a la célula bacteriana mientras la cápside viral permanece en el exterior.
- El despojo es el proceso en que la cápside vírica es degradada por enzimas virales o del huésped, liberando así el ácido nucleico y las proteínas del genoma vírico los cuales comenzaran a replicarse.
- La replicación implica la síntesis de ARN mensajero (ARNm) vírico en todos los virus con rasgos de ARN positivos, la síntesis de proteínas víricas, el ensamblaje de proteínas víricas y la replicación del genoma viral. El proceso de replicación es esencial para mantener la estabilidad de la información genética contenida en el ADN. Esta replicación utiliza enzimas idénticas a las involucradas en la replicación del ADN celular y una característica común es la presencia de estructuras circulares temporales por lo menos en algunas de dicho proceso.
- Tras el ensamblaje de partículas víricas, a menudo se produce una modificación postraduccional de las proteínas víricas. En virus como el VIH, esta modificación (a veces llamada «maduración»), se produce después de que el virus haya sido liberado de la célula huésped. El ensamblaje puede producir nuevas partículas virales. Los virus pueden autoensamblarse en un proceso similar a la cristalización, ya que las partículas virales, al igual que los cristales, constituyen estructuras que se encuentran en un estado mínimo de energía libre. Sin embargo, el genoma viral también puede especificar ciertos factores «morfogenéticos» que no contribuyen directamente a formar la estructura del virión, pero son necesarios para el proceso de ensamblaje. El fenómeno de autoensamblaje ocurre en la formación de diversas estructuras biológicas.
- Los virus son liberados de la célula huésped por lisis, un proceso que mata a la célula reventando su membrana. Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Efectos en la célula huésped
La variedad de efectos estructurales y bioquímicos de los virus sobre las células huésped es grande. Reciben el nombre de «efectos citopáticos». La mayoría de infecciones víricas acaban provocando la muerte de la célula huésped, entre cuyas causas están la lisis de la célula, las alteraciones de la membrana superficial de la célula y la apoptosis. A menudo, la muerte de la célula es causada por el paro de sus actividades normales debido a la supresión por proteínas específicas del virus, que no son todas componentes de la partícula vírica.
Algunos virus no causan cambios aparentes en la célula infectada. Las células en que los virus está latente e inactivo presentan pocos signos de infección y a menudo funcionan normalmente. Esto causa infecciones persistentes y el virus a menudo permanece durmiente durante muchos meses o años. Este suele ser el caso del herpes simple. Algunos virus, como el virus de Epstein-Barr, a menudo hacen proliferar las células sin causar malignidad, pero otros, como los papilomavirus, son una causa demostrada de cáncer.
Fuente: Wikipedia